research interests
| <<< back | research interests |
|
|
My research lies at the interface between physics and biology and merges a theoretical approach with numerical simulations and small-scale experiments.
I am particularly interested in locomotion and fascinated by the emergence of cooperation among microorganisms. The paradigm of unicellularity as an individualistic form of life has been undermined by the discovery of communication and cooperation phenomena across the tree of life. Cooperation offers clever ways to elude the physical constraints to locomotion of single cells. For instance, isolated fungal spores forcibly ejected into the air strive against an enormous resistance due to their microscopic size; but hundreds of thousands of spores are able to travel tens of times further by synchronizing their ejection. Bacterial biofilms are able to exert forces and colonize surfaces even when single cells would be unable to translocate individually. In these systems, I use biomechanics as a tool to understand locomotion as a biological strategy.
|
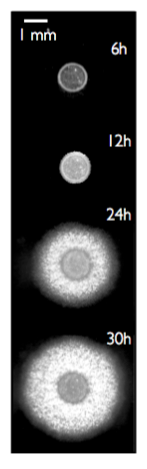
Bacterial biofilms are heterogeneous colonies of differentiated bacteria that live in association with surfaces and present
a remarkable spatio-temporal organization.
The formation of a mature biofilm begins from cell attachment to a surface and proliferation. External signals arising
from threatening events like starvation, trigger an irreversible commitment to the surface.
The hallmark of biofilm formation is the simultaneous loss of flagella mediated motility and production
of an extracellular polymeric matrix encasing the cells and holding the community together. In the Gram-positive
soil bacterium Bacillus subtilis, most cells across the biofilm eventually give up their individual motility.
However, a slower kind of surface translocation arises that is not mediated by flagella and enables the biofilm
to colonize the surface and increase nutrient uptake. In order to spread the biofilm needs to overcome friction
with the surface: I am interested in understanding what are the physical forces that the biofilm exerts and what
is their relation with the spatiotemporal patterns of gene expression. >>> |
 |
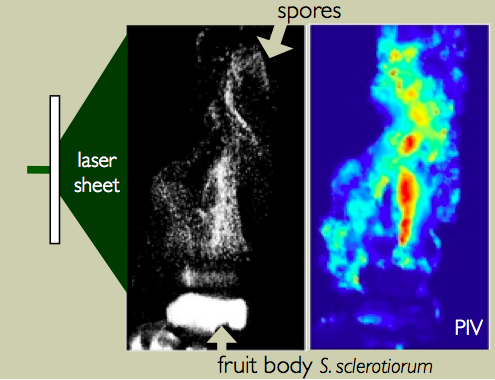
Microscopic spores allow ascomycete fungi to disperse between ephemeral habitats. Spore range is an important factor in a species evolutionary success, since the probability of spore germination depends on the environment where it eventually lands. Although no range of dispersal can guarantee success, there is at least one important constraint that spores need to fullfill in order to germinate: they need not fall back on the parent fungus. This might seem quite an easy task, especially if we consider that spores are launched at enormous velocities that range from 1 m/s to even 20 m/s. |
However, at the micron scale air drag impacts severely on the dynamics of particle motion and spores are decelerated to rest after travelling only few mm. This ballistic range is not large enough to cross the boundary layer of still air that surrounds the fruit body. For species that rely on wind to disperse their propagules this is a major obstacle, because spores that do not cross the boundary layer cannot be entrained by the dispersive air flows and eventually fall back on the parent fungus. I am interested in the disparate ways fungi have evolved to get around this constraint.
Refs: M Roper*, A Seminara*, MM Bandi, A Cobb, H Dillard, A Pringle, PNAS 2010 J Fritz, A Seminara, M Roper, A Pringle and MP Brenner, Proc. Roc. Soc. Interface 2013 F Trail and A Seminara, in review. >>> more details |
| <<< back | PhD research projects |
|
My background is in theoretical physics; during my PhD I worked on various aspects of turbulent transport related to the properties of passively transported fields and to the fate of cloud droplets growing by condensation in a turbulent environment. |
As a part of my PhD I have been working on the problem of condensation/evaporation of microdroplets
in turbulent moist environment. This general issue is relevant for the understanding of
rain-initiation efficiency in warm clouds and for technological issues like the optimization of
spark-ignition engines. Turbulent transport is found to play a crucial role in that it provides
strong fluctuations for droplet growth. Due to correlations between droplets and vapor field,
mean-field type arguments are found to be inadequate to capture relevant aspects of the
problem.
|  |
Refs: |
The statistical properties of systems in thermal equilibrium are fully characterized by the
knowledge of the integrals of motion. Turbulence is the paradigm of systems with many interacting degrees of freedom, strongly out of equilibrium. Here the integrals of motion are insufficient to describe the statistics, as they cannot capture the distinctive features of turbulence: intermittency and anomalous scaling. Instead, these have been brilliantly traced back to statistical integrals of motion whose nature strongly differs from the dynamical integrals of motion of equilibrium systems. Anomalous scaling in turbulence is observed in correspondence of a flux of the conserved quantities through the whole inertial range of scales showing the non-equilibrium nature of the process. In most cases, no upscale flux is present and the statistics at scales larger than the injection scale is expected to recover thermal equilibrium. Surprisingly enough, this is not true in the general case. |
|
Refs: |
The last part of my PhD was dedicated to the problem of diffusion over rocky shorelines. It turns out that the
observed fractality of a consistent portion of the world coastline can be promoted to conformal
invariance. This allows to investigate the problem of diffusion toward the coastline
using results of harmonic analysis for the potential in the vicinity of a conformally invariant
curve. The flux of a pollutant diffusing toward the shoreline is shown to display spatial
intermittency. >>> |
 |